http://www.collembola.org/publicat/senstaxy.htm
-
Last updated on
2022.01.30
by Frans Janssens
Frans Janssens,
Department of Biology, University of Antwerp, Antwerp, B-2020, Belgium
Introduction
Yosii (1961, cited from André, 1988:503) applied in a systematic way
the topology of the chaetae in its description of Xenylla.
This taxonomic descriptive system is the so-called chaetotaxy.
André (1988:503-527) extended Yosii's original chaetotaxy
into a phanerotaxy, including the topology of all setiform
cuticular organs.
A 'phanère' is any organ arising from the integument
(Séguy, 1967; Grassé, 1975 cited from André, 1988:505).
Massoud & Ellis (1977, cited from André, 1988:505) distinguished
four types of phaneres: setae, scales, trichobothria and setiform sensilla,
characterised by a setiform projection of the cuticula articulating with the
body wall by a membraneous socket so it is free to move.
André (1988:520-521) already pointed out that the differentiation
between setae and setiform sensilla is inappropriate, since it suggests that
'normal' setae would have no sensory function.
Scales are highly modified flattened and broadened setae.
Trichobothria are strongly elongated setae with a socket placed in a small
cup-shaped cuticular depression.
In other words, all such phaneres are sensory organs, sensilla.
But the dorsal cephalic epidermal organs include
also several non-setiform types of peripheral sensory organs,
such as the post antennal organ and the ocelli.
In this paper, we extend André's phanerotaxy to include also
those non-setiform sensory organs.
External cuticular sensory organs
External sensory organs are innervated by the dendrite(s)
of one or more bipolar neurons.
The cell body of the bipolar neuron lies beneath the epidermis;
their dendrite(s) penetrate the epidermis and terminate in conjunction with
the cuticula.
The neuron is enclosed by several accessory cells.
Accessory cells are modified epidermal cells which form concentric sheats
around the dendrites and which build the stimulus receptor
external structure.
In trichoid sensilla,
the outer accessory cells form the socket (tormogen cell)
and the shaft (trichogen cell) of the sensory organ.
The inner cell (thecogen cell) secretes a cuticula-like matrix around the tip
of the dendrite(s), called the dendritic cap.
The cephalic external sensory organs are located within the epidermis
and can be classified in three types
of receptors (classification criterion = type of stimulus):
mechanoreceptors,
photoreceptors, and
chemoreceptors.
The mechanoreceptors are typically trichoid sensilla in the form of
setae (chaetae), scales or trichobothria.
They detect touch and air movement
(and possibly sound,
as sound is a periodic low frequency modulated air movement).
Basic structure: a trichogen cell produces a non-perforated seta,
a tormogen cell produces the socket,
a neuron with its dendrite in the seta and its axon to the cerebron.
 |
Fig.phr. Left ocular area
Entomobrya intermedia
2007.11.01 © Valentine, B.
The photoreceptors are typically dome-shaped cuticular structures.
The composed eye of Collembola has maximum 8 single eyes (?ocelli?,?omma?).
They are arranged in a dedicated pattern.
Each single eye has been designated a unique name (A to H)
(see fig. phr.: the ocular area of Entomobrya intermedia).
Basic structure:
four Semper cells have both corneogeneous and crystallogeneous functions:
they produce apically the dome-shaped corneula,
proximally they secrete the crystalline cone
(Barra, 1971:346-351).
The chemoreceptors are in the form of trichoid sensilla and
in the form of a dedicated
post antennal organ (of which the chemoreceptor function is currently not
certain but assumed).
Such olfactory sensors detect airborne molecules.
Basic structure of a trichoid chemoreceptor:
a trichogen cell produces a perforated seta,
a tormogen cell produces the socket,
a neuron with its dendrite in the seta and its axon to the cerebron.
Basic structure of the post antennal organ: large epidermal glial cells
envelope the inner part of the dendrite of a neuron in the cerebron,
an inner glial cell forms the perforated PAO vesicles
(that contain the outer part of the dendrites of the neuron),
2? outer glial cells form the cuticular depression of the PAO (?socket?),
a basal glial cell is apically associated with the neuron in the cerebron
(Karhuize, 1971 cited from Palissa, 2000:12-14).
The glial cells are always present, even when the PAO is externally absent
(Becker, 1910 cited from Palissa, 2000:13-14).
In Symphypleona, the PAO is replaced by a non-homolog post antennal
trichoid sensillum
(Becker, 1910 cited from Palissa, 2000:14).
Basic structure: an epidermal neuron, enveloped by 2 inner glial cells
and 1 large outer glial cell (that has an exosecretion function)
(Altner, Ernst & Karuhize, 1970 cited from Palissa, 2000:14).
Designation of external sensory organs
In accordance with the regression of the prothorax,
the prothoracic chaetotaxy has been simplified (Cassagnau, 1974:302-303).
The dorsal chaetotaxy of the thoracic segments basically consists
of three rows of setae, 'a', 'm' and 'p', as proposed by Yosii (1956)
(André, 1988:514).
The naming of the last two cephalic rows of setae [in Xenylla] of da Gama,
namely cervicals (c) and parietals (p) is not generally followed.
Yosii (1956) proposed to name the most posterior row of the dorsal
cephalic setae, 'c', and the row immediately before, 'p'.
In 1960, Yosii changed his mind and reversed the two namings.
da Gama [(1980)] still follows the former terminology
while Cassagnau (1975) and others have adopted the latter
(André, 1988:508).
Thibaud (1970:134,142,144) in Ceratophysella bengtssoni (fig.10A,B),
in Schaefferia coeca (fig.11A,B), and
in Typhlogastrura balazuci (fig.12A,B)
describes the posterior region of the head as c-row and p-row setae
with respectively 4 and 7 setae,
and refers to the protergal setae as the medial row.
Cassagnau (1974:301[fig.1],302,303) describes
in his model of the cephalic chaetotaxy of the Poduromorpha
the occipital region of the head as c-row and p-row setae,
with respectively 4 and 5 setae,
and refers to the protergal setae as the anterior row or
posterior row, which he considers more evolutionary stable
than the medial row (1974:304).
André (1988:509[fig.2B],514[fig.6A]) describes
in the phanerotaxy of Xenylla jocquei
the posterior region of the head as c-row and p-row setae
and refers to the protergal setae as the anterior row.
Jordana & Arbea (1997:62[fig.23]) in Hypogastruridae
follow the original terminology of c-row and p-row of setae
at the posterior region of the head
and refer to the protergal setae as the medial row,
which is in-line with Yosii (Cassagnau 1974:304).
Fjellberg (1998:15,30[fig.10H]) describes
in the chaetotaxy of the Poduromorpha
the posterior region of the head as c-row and p-row setae
and refers to the protergal setae as the posterior row.
Fig.cc. Nomenclature of cephalic setae in basal Hypogastruridae (Modified after Cassagnau (1974))
Janssens, F. 2004.
 |
We define an alternative schema for the setal pattern on these regions
that is derived from Cassagnau (1974:301)
but takes into account the cephalic chaetotactic model of Thibaud (1970)
and the protergal chaetotactic model of Fjellberg (1998).
In our postoccipito-protergal chaetotactic model,
the poduromorphan cephalic 'cervical' and 'parietal' rows of setae
and the prothoracic posterior row of setae
are derived from an ancestral protergite
with generalised anterior, medial and posterior row of setae.
A transvers region of desclerotisation of the protergite
inbetween the medial setae and posterior setae subdivided the protergite into
two sclerites: the anteroprotergite and the posteroprotergite.
This secondary dorsolateral membraneous region improved the free
dorsoventral movement of the head capsule relative to the trunk.
The broad anteroprotergite bears the anterior and medial row of setae.
The anteroprotergite fuses with the tergite of the posterior cephalic segment.
The ancestral prothoracic anterior row of setae (a-setae) becomes
the by Yosii (1960) designated cephalic row of cervicals (c-setae).
The ancestral prothoracic medial row of setae (m-setae) becomes
the by Yosii (1960) designated cephalic row of parietals (p-setae).
The narrow posteroprotergite bears
the remaining ancestral posterior row of setae (p-setae),
in-line with Fjellberg's protergal model (1998).
In the hypothetical primitive homochaetose scheme of Cassagnau (1974:305),
each thoracic demi-tergite counts 7 setae in
the a-row and m-row and 6 setae in the p-row; setae m7 and p4 are sensilla.
The cephalic c-row and p-row count only 5 setae.
In our scheme (fig.cc), we redesignate several setae of the c-, p-, g-
and l-rows, in-line with Thibaud (1970) and to reflect our
labioprotergal fusion hypothesis, as such:
setae g1 and g1' of Cassagnau (1974:301) respectively become p6 and p7
of Thibaud (1970,134,142,144);
setae l0 and l1 respectively become c6 and c7; and
seta g2 replaces c5 and vice versa.
In this way, the cephalic p- and c-row have each a complete row of 7 setae,
complying to the ancestral scheme as proposed by Cassagnau (1974:305).
The g-row is reduced to g2-5; the l-row is reduced to l2-l5.
Open issues:
What with seta p7 as sensillum?
Interchangeability of external cuticular sensory organs
It has been observed that ocellus B was replaced by a second post antennal
organ in a teratological specimen of Brachystomella parvula from
Mozambique (Cardoso, 19xx:yy).
In some species with reduced number of ocelli it can be observed that a normal
seta appears at the location of the missing ocellus.
In Friesea in general, and more specifically in Friesea cruchagae it can be
observed that the post antennal organ is missing and at its position
a mesochaeta is found
(Arbea & Jordana in Ramos & al., 1997:291,fig.128A).
In Anurida bisetosa, four ocellar setae are found; apparently,
ocelli D and H are absent and ocellus G has been replaced by a mesochaeta
(Arbea & Jordana in Ramos & al., 1997:392,fig.168A).
In Gastranurida denisi, also four ocellar setae are found; in this species
ocelli A, C, D and E are absent and ocellus H has been replaced by a mesochaeta
(Arbea & Jordana in Ramos & al., 1997:408,fig.175A).
And in Monobella, Protanura, Catalanura, Neanura,
Deutonura and Endonura only ocelli A and G are present
and ocellus B appears to be replaced by a microchaeta
(Arbea & Jordana in Ramos & al., 1997:417-466).
 |
Fig.luc. Right ocular sensotaxy in
Ceratophysella lucifuga.
Modified after
Skarzynski, D. © 2007
Case study.
The eyepatch of Ceratophysella lucifuga (Hypogastruridae) has the
maximum number of eight ocelli in the typical ocellar configuration.
However, some specimens show a variant configuration (fig.luc).
The left specimen represents the usual configuration of ocelli
and ocular setae of the right ocellar area.
In the right specimen, an aberant configuration can be observed.
Contrary to Skarzynski's suggestion (Skarzynski, 2007:276,Fig.1I), that ocellus
G is dislocated in the anterior direction, in our hypothesis, the original
ocular seta oc3 is replaced by an ocellus tentatively designated here as 0C3,
and the original ocellus F is replaced by a seta designated here as f.
This case is a nice illustration of the interchangeability of the ocular
sensilla.
All receptors are structurally part of the integument and are made by
modified epidermal cells. In principle, they are interchangeable.
As a rule, neurons and accessory cells of a given sensillum are derivatives
of the same sensory organ progenitor cell.
Depending on the fate of the progenitor cell, it will build a
mechanoreceptive,
photoreceptive, or
chemoreceptive sensillum.
Cephalic sensotactic raster of cuticular sensory organs in Hypogastruridae
We have used a radial concentric topological grid,
formed by the virtual intersection of seven paracoronal planes of
section and ten meridians (see fig.x),
to describe the position of the cephalic cuticular sensory organs.
The seven paracoronal planes of section map to the
seven setae of the p-row sensu Thibaud (1970:134,142,144).
The meridians rotate around the post-occiputal ventro-dorsal axis:
the arbitrary center of the head.
| Meridian | Paracoronal |
|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|
| 0 | - | - | - | - | - | - | a0 |
|---|
| 1 | v1 | d1 | d2 | d3 | d4 | d5 | - |
|---|
| 2 | v2 | sd1 | sd2 | sd3 | sd4 | sd5 | - |
|---|
| 3 | - | oc1 | oc2 | H | C | B | PAO |
|---|
| 4 | - | G | - | oc3 | D | A | ab |
|---|
| 5 | - | - | F | oc4 | E | g5 | l5 |
|---|
| 6 | - | - | - | - | - | g4 | l4 |
|---|
| 7 | - | - | - | - | - | g3 | l3 |
|---|
| 8 | - | - | - | - | - | g2=c5 | l2 |
|---|
| 9 | c1 | c2 | c3 | c4 | c5=- | c6=g2 | c7=l1 |
|---|
| 9' | p1 | p2 | p3 | p4 | p5 | p6=g1 | p7=g1' |
|---|
Tab.X. Generalised dorsal cephalic sensotaxy in Hypogastruridae
|
Fig.x. Generalised dorsal cephalic sensotaxy in Hypogastruridae
Janssens, F. 2007.
 |
|
Although the ocellar seta oc4 was not available in the orginal cephalic
chaetotaxy as prestented by Cassagnau (1974:301),
it has been taken into account in our sensotactic raster in compliance with
the generalised cephalic chaetotaxy of Hypogastruridae as described by
Jordana & Arbea (1997:62,fig.23A).
 |
Fig.ab. Anterior part of left ocular area in
Ceratophysella lucifuga.
Modified after
Skarzynski, D. © 2007
In the postantennal organ (PAO) of Ceratophysella lucifuga
(Hypogastruridae), the accessory boss (ab) is present. While the central lobe
of the PAO is located in front of ocellus B, the accessory boss is located in
front of ocellus A (fig.ab).
It is clear that the PAO fills two positions in the 7th paracoronal of the
sensotactic raster: (3,7) and (4,7).
Onychiuridae
Onychiuridae do not have any ocelli. We will show how the ocellar
positions in the sensotactic raster have been occupied by setae.
The table below (table m) maps the general cephalic sensotactic designations
with the generalised designations of the cephalic setae in Onychiurus
and Protaphorura as described in Jordana & al. (1997:479,fig.190).
Setae that replace ocelli are designated by lower case characters of the same
name as the ocellus. E.g.: seta 'b' replaces ocellus 'B'.
Seta designations indicated with an asterisk (*) are deduced since they
were not designated by Jordana & al. (1997:479,fig.190).
| Paracoronal | Meridian 0 | Meridian 1 | Meridian 2 | Meridian 3 | Meridian 4 | Meridian 5 | Meridian 6 | Meridian 7 | Meridian 8 | Meridian 9 | Meridian 9' |
|---|
| 7 | a0 | = sd'5 | = g10 | PAO | | l5 = l6'* | l4 = l5'* | l3 = l4 | l2 = l3 | c7 = g1 | p7 |
|---|
| 6 | d5 = a0' | sd5 | b = sd'4 | a = g9 | g5 = l6* | g4 = l5 | g3 = g5 | g2 = ca6 | c6 = g1' | p6 |
|---|
| 5 | | d4 | sd4 | c = sd3 | d = g8 | e = g7 | | = cm5 | = cb6 | c5 = cp6 | p5 |
|---|
| 4 | | d3 | sd3 = sd3' | h = sd'3 | oc3 = sd'1 | oc4 = g6 | | = cm4 | = cb5 | c4 = cp4 | p4 |
|---|
| 3 | | d2 | sd2 | oc2 = sd'2 | | f = ca5 | | = cm3 | = cb4 | c3 = cp3 | p3 |
|---|
| 2 | | d1 | sd1 | oc1 = v4 | g = v3 | | | = cm2 | = cb2 | c2 = cp2 | p2 |
|---|
| 1 | = d0 | v1 | v2 | | | | | = cm1 | = cb1 | c1 = cp1 | p1 |
|---|
Table m. Mapping of the general sensotaxy with the onychiurid cephalic chaetotaxy (cf. Jordana & al. (1997:479,fig.190))
Janssens, F. 2006.
The setae typical for Onychiuridae and absent in Hypogastruridae
(designations starting with '=') are well fitted into the sensotactic raster.
After mapping the designations of the cephalic chaetotaxy of Onychiuridae,
it is clear that in the ocular patch (meridians 3, 4 and 5) all ocelli
have been replaced by setae.
In Onychiuridae, the posteroprotergite appears to bear more than just
the p-setae (meridians 7, 8, 9 and 9').
Is this a secondarely derived character,
the result of polychaetosis?
Or are the Onychiuridae the most archaic poduromorphans,
and is the primitive chloting polychaetose and not homochaetose?
Are Onychiuridae an early side branch with a different,
larger posteroprotergite?
Open issues:
What with onychiurid setae g2, g3, g4 and l1?
Ocular sensotactic raster of cuticular sensory organs
We will now present some case studies of ocular sensotactic rasters of some
poduromorph and entomobryomorph species.
Poduromorpha
 |
Fig.soc. Left ocular sensotaxy in
Hypogastrura socialis.
Modified after
Schulz, H.-J. & Schubert, H.D. © 2006
Case study 1.
The eyepatch of Hypogastrura socialis (Hypogastruridae) has the
typical and maximum number of ocelli;
the 8 ocelli are designated as A, B, C, D, E, F, G and H (see fig.soc).
The ocellar setae are designated oc1, oc2 and oc3.
The ocellar seta oc4 is missing.
Ocellar setae oc1 and oc3 are mesochaetae.
Ocellar seta oc2 is a macrochaeta.
The ocellar sensotactic raster is not deformed and therefore easily recognised.
 |
Fig.y'. Right ocular sensotaxy in
Ceratophysella denticulata.
Modified after
Hopkin, S.P. © 2005
|
 |
Fig.y. Right ocular sensotaxy in
Tijucameria mame.
Modified after
de Mendonça, M.C. & Fernandes, L.H. 2005:700(fig.3).
|
Case study 2.
Comparing the sensilla of the ocular area of
Ceratophysella denticulata (Hypogastruridae)
(specimen collected from Swanley, Kent, UK in May 1950;
micrograph by Hopkin, S.P. 2005)
and of Tijucameria mame
de Mendonça, M.C. & Fernandes, L.H. 2005:699-701
(Neanuridae, Pseudachorutinae)
might serve as an example to illustrate
the functional interreplacebility of the sensillar positions
in the sensotactic scheme.
The eyepatch of Ceratophysella denticulata has the typical
and maximum number of ocelli;
the 8 ocelli are designated as A, B, C, D, E, F, G and H (see fig.y').
The ocellar setae are designated oc1, oc2 and oc3.
The ocellar seta oc4 is missing.
Ocellar setae oc1 and oc3 are mesochaetae.
Ocellar seta oc2 is a macrochaeta.
The eyepatch of Tijucameria mame has a reduced number of ocelli;
the 6 ocelli are designated as A, B, C, E, F and G (see fig.y).
In Tijucameria mame,
the two missing ocelli, designated as D and H, are replaced by two microchaetae,
here designated as setae d and h.
The ocellar setae oc1, oc2, oc3 and oc4 are present as microchaetae.
The sensotactic raster of the eyepatch of Tijucameria mame
is quite deformed.
Compared to the raster of the generalised poduromorph scheme:
1. it is contracted radio-transversally at the level of the fourth
paracoronal plane of section, reducing the distance inbetween setae h-oc3-oc4;
more specifically, setae h and oc4 are approaching the center of the eyepatch:
seta oc3;
2. it is widened transversally at the level of the fifth
paracoronal plane of section, enlarging the distance inbetween C-d-E.
The deformation of the sensotactic raster results mainly in strongly
deformed third and fifth meridians (fig.y). To avoid overloading the figure,
only the meridians of the sensotactic raster are drawn.
Case study 3.
The ocular sensotactic raster is situated on the meridians 3 to 5
and paracoronals 2 to 6. Each raster position can be identified by a coordinate
specifying the number of the meridian and paracoronal as such : (m,p).
As an example: the raster position of ocellus A is (4,6).
| Species | Raster position (meridian,paracoronal) |
|---|
| (3,2) | (3,3) | (3,4) | (3,5) | (3,6) | (4,2) | (4,3) | (4,4) | (4,5) | (4,6) | (5,2) | (5,3) | (5,4) | (5,5) |
|---|
| Body plan | Oc1 | Oc2 | H | C | B | G | - | Oc3 | D | A | - | F | Oc4 | E |
| chiangdaoensis | M | M | me | C | - | G | - | - | - | A | - | - | - | - |
| colombiana | - | M | me | - | - | G | - | - | - | A | - | - | - | - |
| meo | - | M | me | - | - | G | - | - | - | A | - | - | - | - |
| modesta | me | M | me | - | - | G | - | - | - | A | - | - | - | - |
| najtae | M | M | me | - | - | G | - | - | D | A | - | - | - | - |
| tapiata | - | me | me | - | - | G | - | - | - | A | - | - | - | - |
| tibiotarsalis | me | M | me | - | - | G | - | - | - | A | - | - | - | - |
Table r. The ocular sensotactic raster in some species of Paranura.
Synapomorphies in the following species of Paranura:
chiangdaoensis, meo, modesta, and tibiotarsalis
(Deharveng, 1989),
najtae
(Deharveng & Weiner, 1984), and
colombiana, tapiata
(Palacios-Vargas & Peñaranda-Parada, 2005),
are the presence of a receptor at raster
positions (3,3), (3,4), (4,2) and (4,6)
and the abscence of a receptor at raster positions (3,6), (4,4), (5,3),
(5,4) and (5,5).
A receptor can be absent or present at raster positions (3,2), (3,5) and (4,5).
The following receptors are always present: ocelli A and G, and the
ocular seta Oc2 (in Paranura context called Ocm).
The latter is mostly a macrochaeta (M), exceptionally a mesochaeta (me).
The optional ocular seta Oc1 (in Paranura context called Ocp)
appears in some species as macrochaeta and in others as mesochaeta.
In the context of this paper, raster position (3,4) is most interesting.
In the generic body plan of the Poduromorpha ocellus H is found at this
position. In Paranura, a mesochaeta (called Oca) is found at this
postion.
 |
Fig.sg. Right ocular sensotaxy in
Stachorutes gracilis.
Modified after
Smolis, A. & Shvejonkova, J.B. © 2006
Case study 4.
The ocular area of Stachorutes gracilis Smolis & Shvejonkova, 2006
shows a reduced number of ocelli and increased number of ocular setae (fig. sg).
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B-PAO) is complete.
However, ocellus H has been substituted by mesoseta h.
Of the meridian 4 pattern (G-+-oc3-D-A), ocellus G is present,
ocellar seta oc3 is absent,
ocellus D is absent and not substituted,
and ocellus A has been replaced by mesoseta a.
Of the meridian 5 pattern (+-F-oc4-E),
ocellus F is present,
ocellar seta oc4 is present,
ocellus E is absent and not substituted.
Absent and not substituted ocelli are indicated in red color.
To conclude: of the four absent ocelli, 2 have been substituted by mesosetae.
 |
Fig.ip. Right ocular sensotaxy in
Israelimeria poliakovae.
Modified after
Weiner, W.M. & Kaprus, I.J. © 2005
Case study 5.
The right ocular area of Israelimeria poliakovae Weiner & Kaprus, 2005
shows a reduced number of ocelli and increased number of ocular setae (fig. ip).
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B-PAO) is complete.
However, ocelli H and C have been substituted by mesosetae h and c.
And the PAO is substituted by mesoseta pao.
Of the meridian 4 pattern (G-+-oc3-D-A), ocelli G and A are present,
ocellar seta oc3 is present,
ocellus D is substituted by mesoseta d.
Of the meridian 5 pattern (+-F-oc4-E),
ocellus F is present,
ocellar seta oc4 is present,
ocellus E is substituted by mesoseta e.
To conclude: all absent ocelli as well as the PAO have been substituted
by mesosetae.
 |
Fig.gh. Left ocular sensotaxy in
Gomphiocephalus hodgsoni.
Modified after
Meyer-Rochow, V.B. & al © 2005
 |
Fig.gh2. Left ocular sensotaxy in
Gomphiocephalus hodgsoni.
Modified after
Meyer-Rochow, V.B. & al © 2005
Case study 6.
The left ocular area of Gomphiocephalus hodgsoni Carpenter, 1908
shows a complete number of ocelli and almost complete number of ocular setae (fig. gh and gh2).
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B-PAO) is complete.
The meridian 4 pattern (G-+-oc3-D-A) is complete.
Of the meridian 5 pattern (+-F-oc4-E),
ocelli F and E are present,
ocellar seta oc4 is absent,
To conclude: all ocelli are present, ocellar setae oc1-3 are present as mesosetae,
ocellar seta oc4 is absent.
Entomobryomorpha
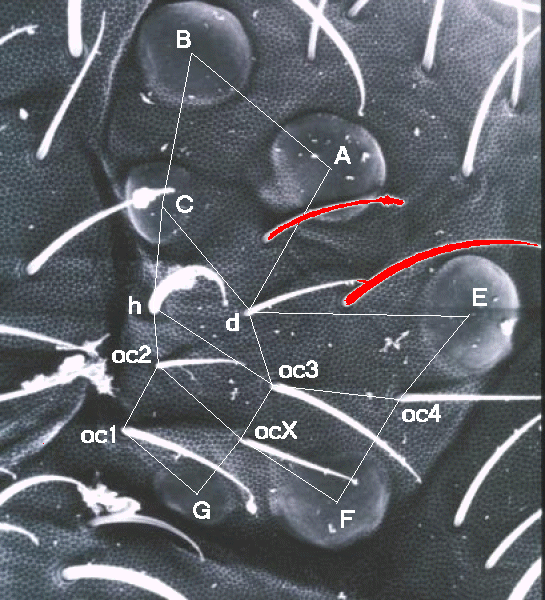 |
Fig.ey. Right ocular sensotaxy in an undefined
glacier active isotomid from Switzerland.
Modified after
Nielsen, L., © 2005
Case study 1: Isotomidae.
The ocular area of an undefined glacier active isotomid from Switzerland
shows a reduced number of ocelli and increased number of ocular setae (fig.ey).
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B) is complete.
Ocellus C is reduced in size,
and ocellus h has been substituted by macroseta h.
Also the meridian 4 pattern (G-ocX-oc3-D-A) is complete.
Ocellus G is reduced in size,
and ocellus D is absent and substituted by mesoseta d.
Note that in this species, the open position in the ocular sensotactic raster
(in the center of oc2, oc3, G, and F) is taken by a seta that we have
designated ocX.
Even the meridian 5 pattern (+-F-oc4-E) is complete.
The two additional setae (red coloured in fig.ey) are
invaginated setae, most probably setae g5 and l5 respectively.
Due to this invagination, the raster is deformed at the level of the
paracoronals 4 and 5.
To conclude: in this isotomid, the two absent ocelli D and H have been
substituted by setae d and h. The ocular sensotactic raster in Isotomidae appears
more complete compared to the raster in Poduromorpha, due the presence of
seta ocX.
In this particular case, the raster is quite deformed due to the invagination
of genal and lateral setae.
Note, that in this particular case,
the substitutions of ocelli by seta in the ocular sensotactic raster
resemble those of Orchesella.
Although quite deformed by the invagination of setae from neighbourhood
cephalic areas, the
ocular sensotactic raster appears to be complete in Isotomidae:
both ocular setae ocX and oc4 are present.
Case study 2: Entomobryidae: Orchesellinae.
The ocular area of Orchesella
shows a reduced number of ocelli and increased number of ocular setae (fig. ldv).
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B) is complete.
Ocellus C is reduced in size,
and ocellus H has been substituted by macroseta h (note that only the setal
socket is visible in the SEM photograph (fig.ldv)).
The meridian 4 pattern (G-+-oc3-D-A) is almost complete.
OcX is absent.
Ocellus G is reduced in size,
and ocellus D is absent and substituted by microseta d.
Even the meridian 5 pattern (+-F-oc4-E) is complete.
Note the the position of oc4 is shifted inwards of the ocullar area
due to the spherical shape of the ocular area surface.
To conclude: in Orchesella, the two absent ocelli have been
substituted by setae.
 |
Fig.e. Right ocular sensotaxy in
Orchesellides carolinae.
Modified after
Jordana & Baquero, 2006:86 Fig.2A
Case study 3: Entomobryidae: Orchesellinae.
The lower half of the ocular sensotactic raster is quite contracted in
Orchesellides carolinae Jordana & Baquero, 2006 (fig.e).
The meridians are indicated in red color, while the paracoronals are
indicated in blue color.
Due to the reduction of ocelli G and H, the lower part of the ocular area
is quite contracted due to which the ocular setae 2, 3 and 4
approach eachother up to such extent that the meridians 3 and 4
are crossing at the level of paracoronal 3.
Note that the headcapsule of Entomobryomorpha is more spherical than
in Poduromorpha having a more dorsoventral flattened headcapsule.
Due to the projection on a flat surface of the spherical constellation of
the ocular sensotactic raster it seems more radially deformed with respect
to that of Poduromorpha.
 |
Fig.r. Right ocular sensotaxy in
Rhynchocyrtus klausi.
Modified after de Mendonça & Fernandes, 2007:48 Fig.9
Case study 4: Entomobryidae: Lepidocyrtinae.
In Rhynchocyrtus klausi from Brazil (fig.r),
the meridian 3 pattern of sensilla (oc1-oc2-H-C-B) is complete.
Ocellus H is reduced in size.
Also the meridian 4 pattern (G-ocX-oc3-D-A) is complete.
Ocellus G is reduced in size.
Note that also in this species, as in Isotomidae,
the open position in the ocular
sensotactic raster (in the center of oc2, oc3, G, and F) is taken by a seta
that we have designated ocX.
Also the meridian 5 pattern (+-F-oc4-E) is complete.
The ocular sensotactic raster appears, as in Isotomidae, more complete
compared to the raster in Poduromorpha, due the presence of seta ocX.
 |
Fig.f. Right ocular sensotaxy in
Lepidocyrtus flexicollis.
Modified after Mateos, E. 2008:40 Fig.5
 |
Fig.c. Right ocular sensotaxy in
Lepidocyrtus curvicollis.
Modified after Mateos, E. 2008:45 Fig.13
Designation of intraocular setae in Lepidocyrtus,
after Mateos (2008): oc1=t, oc2=s, oc3=q, oc4=p, ocX=scale(s).
In Lepidocyrtus flexicollis, oc3 is absent, ocX is present in
the form of a scale.
In Lepidocyrtus curvicollis, oc3 is present, ocX is present in
the form of two scales.
 |
Fig.s1. Right ocular sensotaxy in
Seira dowlingi, 1st instar.
Modified after Soto-Adames, 2008:10 Fig.24
 |
Fig.s2. Right ocular sensotaxy in
Seira dowlingi, adult.
Modified after Soto-Adames, 2008:12 Fig.27
Case study 5: Entomobryidae: Seirinae.
In the first instar of Seira dowlingi from Puerto Rico (fig.s1),
the meridian 3 pattern of sensilla (oc1-oc2-x-C-B) is incomplete.
Ocellus H is missing (missing sensilla are indicated with x).
Also the meridian 4 pattern (x-x-oc3-D-A) is incomplete.
Ocellus G and ocular seta ocX is missing.
Also the meridian 5 pattern (+-F-x-E) is incomplete.
Ocular seta oc4 is missing.
Given the missing ocular setae oc4 and ocX, the ocular sensotactic raster
of the first instar of Seira
appears to match with that of Hypogastrura and Ceratophysella.
In the adult instar of Seira dowlingi from Puerto Rico (fig.s2),
the meridian 3 pattern of sensilla (oc1-oc2-H-C-B) is complete.
Also the meridian 4 pattern (G-ocX-oc3-D-A) is complete.
Also the meridian 5 pattern (+-F-oc4-E) is complete.
Given the presence of both ocular setae oc4 and ocX, the ocular sensotactic raster
of the adult instar of Seira is more complete as in Lepidocyrtinae.
Most adults have five setae in the eyepatch, probably homologous to setae
p, q, r, v, and t in Lepidocyrtus (Mari Mutt 1986).
Note: q=oc3, p=oc4, r=ocX, v=oc2, t=oc1.
Setae oc3(=q), oc2(=v), and oc1(=t) are primary (Fig. s1).
Additional setae first appear during the 4th instar, but are unstable:
of seven specimens examined four have only the primary setae,
one specimen adds ocX, but two others add oc4 instead.
All ocular setae are present in all specimens beginning at 5th istar.
(modified after Soto-Adames, 2007:7).
 |
Fig.tom. Left ocular sensotaxy
Tomocerus sp.
2001 © Plant, N.
Case study 6: Tomoceridae.
The ocular area of a Tomocerus sp. from the USA
shows a reduced number of ocelli and increased number of ocular setae (fig.tom).
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B) is complete.
But ocellus H has been substituted by mesoseta h.
Note that in fig.tom oc1 en oc2 are only represented by sockets.
The meridian 4 pattern (G-+-oc3-D-A) is almost complete.
OcX is absent.
0c3 is a macroseta.
The meridian 5 pattern (+-F-+-E) is incomplete.
Ocellus E has been substituted by mesoseta e.
The ocellar seta oc4 is missing.
 |
Fig.trog. Left ocular sensotaxy
Trogolaphysa nigrolineata first instar
Modified after 2022 © Cipola, N.G. & al. Fif.19A
Case study 7: Paronellidae.
In the ocular area of the first instar of Trogolaphysa nigrolineata
from Brazil
the ocular setae p, r and t (fig.trog)
correspond respectively with ocular setae oc3, oc2 and oc1 of Poduromorpha.
The meridian 3 pattern of sensilla (r=oc1-t=oc2-H-C-B) is complete.
The meridian 4 pattern (G-+-p=oc3-D-A) is complete.
OcX is absent.
The meridian 5 pattern (+-F-+-E) is incomplete.
Ocellar seta oc4 inbetween ocelli E and F
is missing as in Poduromorpha.
Seta g5 is present.
Ocelli E and F are displaced.
| Family | ocX | oc4 |
|---|
| Tomoceridae | absent | absent |
| Isotomidae | present | present |
| Orchesellidae | absent | present |
| Entomobryidae: Lepidocyrtinae | present | present |
| Entomobryidae: Seirinae (1st instar) | absent | absent |
| Entomobryidae: Seirinae (adult) | present | present |
| Paronellidae: (1st instar) | absent | absent |
Tab.Ent. Entomobryomorpha. Summary
Symphypleona
 |
Fig.Ba. Right ocular sensotaxy in
Borgesminthurinus andinus from Bolivia.
Modified after
Palacios-Vargas JG & Vacaflores-Argandoña MR, © 2020 Fig.27
Case study 1: Borgesminthurinus.
The ocular sensotaxy of Borgesminthurinus andinus from Bolivia
matches remarkably well that of Poduromorpha.
The meridian 3 pattern of sensilla (oc1-oc2-H-C-B) is complete.
Ocellus C is reduced in size,
Ocellar seta ocX (of Entomobryomorpha) inbetween ocellus G and ocular seta oc3
is missing in the meridian 4 pattern (G-+-oc3-D-A) as in Poduromorpha.
Ocellus D is reduced in size,
Ocellar seta oc4 inbetween ocelli E and F of the meridian 5 pattern
is missing (+-F-+-E) as in Poduromorpha.
To conclude:
The ocular sensotactic raster in Borgesminthurinus appears
to match that of Poduromorpha,
suggesting a direct ancestral relationship
between Symphypleona and Poduromorpha.
To be completed...
Bibliography


